Развитие нервной системы
Нервная система делится на центральную и периферическую. В периферическую нервную систему входят корешки, сплетения и нервы. ЦНС состоит из головного и спинного мозга. Изучение онтогенеза ЦНС позволило установить, что головной мозг образуется из мозговых пузырей, возникающих в результате неравномерного роста передних отделов медуллярной трубки. Из этих пузырей формируются передний мозг, средний мозг и ромбовидный мозг. В дальнейшем из переднего мозга образуются конечный и промежуточный мозг, а ромбовидный мозг также разделяется соответственно на задний и продолговатый мозг.
Из конечного мозга соответственно формируются полушария большого мозга, базальные ганглии, из промежуточного мозга – таламус, эпиталамус, гипоталамус, метаталамус, зрительные тракты и нервы, сетчатка. Зрительные нервы и сетчатка являются отделами ЦНС, как бы вынесенными за пределы головного мозга. Из среднего мозга образуются пластинка четверохолмия и ножки мозга. Из заднего мозга формируются мост и мозжечок. Мост мозга граничит внизу с продолговатым мозгом. Задняя часть медуллярной трубки формирует спинной мозг, а ее полость превращается в центральный канал спинного мозга. В конечном мозге располагаются боковые желудочки, в промежуточном мозге – III желудочек, в среднем мозге – водопровод мозга, соединяющий III и IV желудочки; IV желудочек находится в заднем и продолговатом мозге.
Морфология нервной клетки
Основу нервной системы составляют нервные клетки. Кроме нервных клеток в нервной системе имеются глиальные клетки и элементы соединительной ткани.
Структура нервных клеток различна. Существуют многочисленные классификации нервных клеток, основанные на форме их тела, протяженности и форме дендритов и других признаках.
По функциональному значению нервные клетки подразделяются на двигательные (моторные), чувствительные (сенсорные) и интернейроны.
Нервная клетка осуществляет две основные функции: а) специфическую – переработку поступающей на нейрон информации и передачу нервного импульса; б) биосинтетическую, направленную на поддержание своей жизнедеятельности. Это находит выражение и в ультраструктуре нервной клетки. Передача информации от одной нервной клетки к другой, объединение нервных клеток в системы и комплексы различной сложности определяют характерные структуры нервной клетки – аксоны, дендриты и синапсы. Органеллы, связанные с обеспечением энергетического обмена, белоксинтезирующей функцией клетки и др., встречаются в большинстве клеток, в нервных клетках они подчинены выполнению их основных функций – переработке и передаче информации.
Тело нервной клетки на электронно-микроскопических фотографиях представляет собой округлое и овальное образование. В центре клетки (или слегка эксцентрично) располагается ядро. Оно содержит ядрышко и окружено наружной и внутренней ядерными мембранами толщиной около 70 А каждая, разделенных перинуклеарным пространством, размеры которого вариабельны. В кариоплазме распределены глыбки хроматина, которые имеют тенденцию скапливаться у внутренней ядерной мембраны. Количество и распределение хроматина в кариоплазме вариабельны в различных нервных клетках.
В цитоплазме нервных клеток располагаются элементы зернистой и незернистой цитоплазматической сети, полисомы, рибосомы, митохондрии, лизосомы, многопузырчатые тельца и другие органеллы.
Структуру нервной клетки представляют: митохондрии, определяющие ее энергетический обмен; ядро, ядрышко, зернистая и незернистая эндоплазматическая сеть, пластинчатый комплекс, полисомы и рибосомы, в основном обеспечивающие белоксинтезирующую функцию клетки; лизосомы и фагосомы – основные органеллы «внутриклеточного пищеварительного тракта»; аксоны, дендриты и синапсы, обеспечивающие морфофункциональную связь отдельных клеток. Полиморфизм строения клеток определяется различной ролью отдельных нейронов в системной деятельности мозга в целом.
Понять структурно-функциональную организацию мозга в целом не представляется возможным без анализа распределения дендритов, аксонов и межнейрональных связей.
Дендриты и их разветвления определяют рецептивное поле той или иной клетки. Они очень вариабельны по форме, величине, разветвленное и ультраструктуре. Обычно от тела клетки отходит несколько дендритов. Количество дендритов, форма их отхождения от нейрона, распределение их ветвей являются определяющими в основанных на методах серебрения классификациях нейронов.
При электронно-микроскопическом исследовании обнаруживается, что тело нервных клеток постепенно переходит в дендрит, резкой границы и выраженных различий в ультраструктуре сомы нейрона и начального отдела крупного дендрита не наблюдается.
Аксоны, так же как и дендриты, играют важнейшую роль в структурно-функциональной организации мозга и механизмах системной его деятельности. Как правило, от тела нервной клетки отходит один аксон, который затем может отдавать многочисленные ветви.
Аксоны покрываются миелиновой оболочкой, образуя миелиновые волокна. Пучки волокон (в которых могут быть отдельные немиелинизированные волокна) составляют белое вещество мозга, черепные и периферические нервы.
При переходе аксона в пресинаптическое окончание, наполненное синаптическими пузырьками, аксон образует обычно колбовидное расширение.
Переплетения аксонов, дендритов и отростков глиальных клеток создают сложные, неповторяющиеся картины нейропиля. Однако именно распределение аксонов и дендритов, их взаиморасположение, афферентно-эфферентные взаимоотношения, закономерности синапсоархитектоники являются определяющим в механизмах замыкательной и интегративной функций мозга.
Взаимосвязи между нервными клетками осуществляются межнейрональными контактами, или синапсами. Синапсы делятся на аксосоматические, образованные аксоном с телом нервной клетки, аксодендритические, расположенные между аксоном и дендритом, и аксо-аксональные, находящиеся между двумя аксонами. Значительно реже встречаются дендро-дендритические синапсы, расположенные между дендритами.
В синапсе выделяют пресинаптический отросток, содержащий пресинаптические пузырьки, и постсинаптическую часть (дендрит, тело клетки или аксон). Активная зона синаптического контакта, в которой осуществляются выделение медиатора и передача импульса, характеризуется увеличением электронной плотности пресинаптической и постсинаптической мембран, разделенных синаптической щелью. По механизмам передачи импульса различают синапсы, в которых эта передача осуществляется с помощью медиаторов, и синапсы, в которых передача импульса происходит электрическим путем, без участия медиаторов.
Существенным моментом в синаптической передаче является то, что в разных системах межнейрональных связей используются различные медиаторы. В настоящее время известно около 30 химически активных веществ (ацетилхолин, дофамин, норадреналин, серотонин, ГАМК и др.), которые играют роль в синаптической передаче импульсов от одной нервной клетки к другой.
В последнее время в качестве посредников в синаптической передаче активно изучаются многочисленные нейропептиды, среди которых наибольшее внимание привлекают энкефалины и эндорфины, субстанция Р. Выделение из пресинаптического отростка медиатора или модулятора синаптической передачи теснейшим образом связано со структурой постсинаптической рецептивной мембраны.
Важную роль в межнейрональных связях играет аксональный транспорт. Принцип его заключается в том, что в теле нервной клетки синтезируется ряд ферментов и сложных молекул, которые затем транспортируются по аксону в его концевые отделы – синапсы.
Система аксонального транспорта является тем основным механизмом, который определяет возобновление и запас медиаторов и модуляторов в пресинаптических окончаниях, а также лежит в основе формирования новых отростков, аксонов и дендритов.
Согласно представлениям о пластичности мозга в целом, в мозге происходят два взаимосвязанных процесса: 1) формирование новых отростков и синапсов; 2) деструкция и исчезновение некоторой части существовавших ранее межнейрональных контактов.
Механизмы аксонального транспорта, связанные с ними процессы синаптогенеза и роста тончайших разветвлений аксонов лежат в основе обучения. адаптации, компенсации нарушенных функций. Расстройство аксонального транспорта приводит к деструкции синаптических окончаний и изменению функционирования определенных систем мозга.
Воздействуя рядом лекарственных веществ и биологически активными веществами, можно влиять на метаболизм нейронов, определяющий их аксональный транспорт, стимулируя его и повышая тем самым возможность компенсаторно– восстановительных процессов.
Усиление аксонального транспорта, рост тончайших ответвлений аксонов и синаптогенез играют положительную роль в осуществлении нормальной работы мозга. При патологии эти явления лежат в основе репаративных, компенсаторно-восстановительных процессов.
Кроме механизмов аксонального транспорта биологически активных веществ, которые идут от тела нервной клетки к синапсам, существует так называемый ретроградный аксональный транспорт веществ от синаптических окончаний к телу нервной клетки. Эти вещества необходимы для поддержания нормального метаболизма тел нервных клеток и, кроме того, несут информацию о состоянии их концевых аппаратов.
Нарушение ретроградного аксонального транспорта приводит к изменениям нормальной работы нервных клеток, а в тяжелых случаях – к ретроградной дегенерации нейронов.
Спинной мозг - medulla spinalis
Общая характеристика
Спинной мозг расположен в позвоночном канале. Имеет вид сдавленного в дорсовентральном направлении тяжа, покрытого мозговыми оболочками.
 Рис. 1. Спинной мозг таксы в позвоночном канале.
Рис. 1. Спинной мозг таксы в позвоночном канале.
Топографически спинной мозг подразделен на шейный (С1 – С8),грудной (Th1 – Th13), поясничный (L1 – L7), крестцовый (S1 – S3) и хвостовой (Ca1-Ca5). Передняя граница спинного мозга соответствует краниальному краю дуги атланта, а задняя: у собак достигает краниального края 7 поясничного позвонка, у кошек – третьего (последнего) крестцового позвонка. На всем протяжении спинной мозг у собак имеет два утолщения в местах отхождения нервов к конечностям: шейное (от С6 до Th2), поясничное (от L4 до S2). У кошек кроме шейного утолщения в области С6 и поясничного в области L5 имеется также грудное утолщение в области Th12. После поясничного утолщения спинной мозг резко сужается, образуя спинномозговой конус, переходящий в концевую нить. Начальный отдел концевой нити содержит нервную ткань спинного мозга, представленную эпендимной трубкой или ее расширением - концевым желудочком (продолжение центрального спинномозгового канала), достигающим у собаки L7/S1, у кошки - Ca1. Конечный отдел терминальной нити, оканчивающийся у собаки на уровне Ca1-Ca3, у кошки - на уровне Ca4-Ca6, представлен твердой оболочкой спинного мозга. Спинномозговой конус, терминальная нить и хвостовые нервы образуют «конский хвост».
- а -conus medullaris
- b –концевая нить
- с- подпаутинное пространство спинного мозга
- d- твердая оболочка мозга
- е- тонкая концевая нить твердой оболочки
- f- cavum epidurale эпидуральная полость.
Из-за опережающего роста позвоночного столба, границы сегментов спинного мозга не совпадают с границами позвонков соответствующих разделов. У собаки 3 крестцовый нейросегмент и включающий хвостовые нейросегменты спинномозговой конус лежат в области 6-7 поясничного позвонка, а у кошек - в области крестцовой кости. Спинной мозг кошки имеет длину около 40см и весит 8-9г, небольшой собаки (таксы) – 48см и весит 14г, большой собаки (немецкой овчарки) – 78см и весит 33г. Оболочки спинного мозга. (Рис 2)Твердая мозговая оболочка (dura mater spinalis, s. pachymeninx) - наружная, построена из плотной соединительной ткани. Покрывает спинной мозг и спинномозговые нервы до места выхода их из межпозвоночных отверстий. Прикрепляясь к дужкам атланта, зубу эпистрофея, по краям межпозвоночных отверстий и хвостовым позвонкам твердая спинномозговая оболочка удерживает спинной мозг в подвешенном состоянии на своеобразных растяжках. Между твердой оболочкой и надкостницей позвоночного канала имеется эпидуральное пространство, заполненное жировой тканью и венозным сплетением. Оно предохраняет спинной мозг от механических сотрясений и обеспечивает его подвижность в позвоночном канале. Наличие эпидурального пространства делает возможным проведение анестезии корешков спинномозговых нервов на их пути к межпозвоночным отверстиям. Эпидуральную анестезию проводят у собаки и кошки между 7 поясничным и 1 крестцовым позвонками, между крестцовой костью и 1 хвостовым позвонком, либо между следующими 2-3 хвостовыми позвонками в зависимости от преследуемой цели.
Паутинная оболочка (arachnoidea spinalis) – средняя, построена из рыхлой соединительной ткани, отделена от твердой мозговой оболочки незначительным субдуральным пространством, заполненным тканевой жидкостью (отдельные авторы утверждают об отсутствии у собак и кошек данного пространства).
Мягкая (сосудистая) мозговая оболочка (pia mater spinalis) – внутренняя, состоит из плотной соединительной ткани. В ней проходят кровеносные сосуды, которые, входя в мозговую ткань, обеспечивают прочное соединение мягкой оболочки со спинным мозгом. От мягкой оболочки в каждом сегменте спинного мозга отходят зубовидные связки, которые, прободая паутинную оболочку, прикрепляются к твердой спинномозговой оболочке, подвешивая внутри ее спинной мозг. От паутинной оболочки мягкая оболочка отделена подпаутинным(субарахноидальным) пространством, заполненным спинномозговой жидкостью.
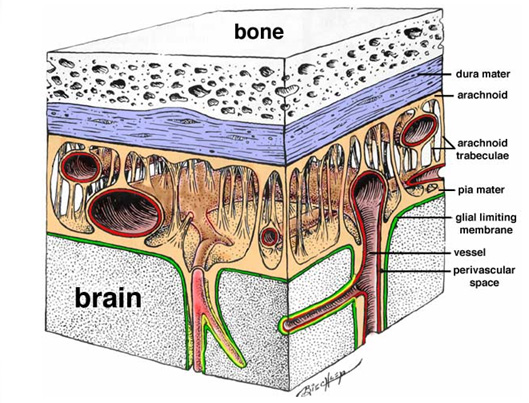
Рис. 2. Оболочки мозга.
Строение спинного мозга
По вентральной поверхности спинного мозга проходят вентральная срединная щель (место расположения центральной спинномозговой артерии и вены) и две латеральные вентральные борозды (место выхода вентральных корешков спинномозговых нервов). По дорсальной поверхности проходят дорсальная срединная борозда и дорсальные латеральные борозды (место вхождения дорсальных корешков спинномозговых нервов).
Спинной мозг состоит из белого мозгового вещества расположенного по периферии и серого мозгового вещества, лежащего в центре. Серое мозговое вещество на поперечном разрезе напоминает очертание буквы Н или крыльев бабочки. Через мостик, соединяющий обе ножки Н-образного серого вещества, серую спайку (центральное промежуточное вещество), проходит центральный канал спинного мозга. На границе спинного и продолговатого мозга центральный канал расширяется и переходит в 4 мозговой желудочек. В области поясничного утолщения спинного мозга центральный канал также расширяется, образуя концевой желудочек,
который в свою очередь, сужаясь, слепо оканчивается в терминальной нити. Форма центрального канала в виде удлиненного овала, высотой у кошек и собак около 100мкм и шириной около 50мкм.
В каждой половине спинного мозга серое вещество залегает в виде дорсального и вентрального столбов, разделенных латеральным и центральным промежуточным веществом. В нижней части дорсальных столбов латерально располагается сетчатое образование, которое представлено идущей поперечно сетью нервных волокон. Оно наиболее выражено в шейном отделе, а наименее - в грудном и поясничном отделах.
В сером веществе спинного мозга локализованы центры, управляющие безусловными рефлексами. На уровне грудных сегментов расположен центр, управляющий мускулатурой позвоночного столба и грудной клетки, на уровне поясничных сегментов лежат центры мускулатуры тазовых конечностей, на уровне последних поясничных сегментов – центры дефекации и мочеиспускания. Морфологически центры представлены ядрами серого мозгового вещества. Ядро формируется телами нервных клеток по принципу единого происхождения, строения и функции. В дорсальном столбе находятся собственное дорсальное ядро (реле в проведении импульсов болевой чувствительности) и грудное ядро (участвует в управлении проприорецептивной чувствительностью от скелетной мускулатуры к мозжечку). В средней части лежат вегетативные ядра: симпатические – в грудопоясничном отделе от C8/Th1 до L4/L5 (промежуточное медиальное ядро) и парасимпатические – в крестцовом отделе от S1 до S3 (промежуточное латеральное ядро). В вентральном столбе - двигательное ядро, от клеток которого отходят соматомоторные волокна. Кроме этих ядер имеются нейроны-переключатели, клетки спаек и ассоциативные клетки (обеспечивают связь между ядрами), канатиковые клетки (образуют своими аксонами пути, соединяющие спинной и головной мозг). Белое мозговое вещество состоит из нервных волокон и формирует проводящие пути. Его больше в краниальной части спинного мозга, а в каудальном направлении количество белого вещества постепенно уменьшается. Столбы серого мозгового вещества делят белое вещество спинного мозга на парные дорсальные, латеральные и вентральные канатики. Располагающееся между дорсальными столбами белое мозговое вещество полностью разделено на 2 половины срединной дорсальной перегородкой. Оба вентральных канатика связывает белая спайка, расположенная вентрально от серой спайки. С дорсальной стороны соответствующая структура отсутствует. В дорсальном канатике проходят восходящие волокна, которые проводят чувствительные импульсы (тактильной и компрессионной чувствительности) без переключения в спинном мозге от периферии к продолговатому мозгу. Волокна из задней части тела, особенно от задней конечности, формируют тонкий пучек (fasciculus cracilis), который по срединной линии примыкает к дорсальной срединной перегородке. Волокна из передней части тела особенно из передних конечностей присоединяются латерально к тонкому пучку, при этом формируя клиновидный пучек (fasciculus cuneatus). Оба пучка на дорсальной поверхности спинного мозга заметны как тяжи, а при переходе в продолговатый мозг объединяются в продолговато-спинномозговой путь (tractus spinobulbaris). В боковом канатике проходят восходящие и нисходящие пути. Восходящие пути располагаются в наружной части канатика и представлены дорсальным спинно-мозжечковым пучком (fasciculus spinocerebralis dorsalis), вентральным спинно-мозжечковым пучком (fasciculus spinocerebralis ventralis), восходящим пучком зрительного бугра и четверохолмия (tractus spinotectothalamicus), который проходит у кошек дорсолатерально и является проводником болевой чувствительности. Нисходящие пути состоят из бокового пучка, выходящего из красного ядра (fasciculus rubrospinalis), вестибулоспинального пучка (fasciculus vestibulospinalis), который лежит вентральнее предыдущего и бокового пирамидного пучка (tractus corticospinalis later-alis s. piramidalis), выраженного у собак и кошек лучше, чем у других домашних животных. Он образован нисходящими волокнами сигмовидной, венечной и эктосильвиевой извилин коры головного мозга и заканчивается на промежуточных нейронах спинного мозга. Лишь незначительная часть волокон у собак и кошек оканчивается на двигательных спинномозговых нейронах. Пересечение у собак и кошек пирамидного пути возле продолговатого мозга незначительно влияет на двигательные функции. Наибольшие изменения возникают при повреждении двигательных центров в коре головного мозга. Вентральный канатик является нисходящим путем и включает вентральный или прямой пирамидный пучек (fasciculus corticospinalis ventralis) и четверохолмный пучек (fasciculus tectospinalis) Восходящие и нисходящие пути не прилегают непосредственно к серому веществу. Узкая полоска белого вещества в виде собственных пучков соединяет восходящие и нисходящие сегменты на одной стороне (ассоциативные клетки) или правую и левую стороны спинного мозга (комиссуральные клетки).
Сосуды спинного мозга
Артерии спинного мозга являются спинномозговыми ветвями позвоночных, межреберных, поясничных и крестцовых артерий. Все эти ветви проникают в позвоночный канал по ходу корешков спинномозговых нервов и образуют на спинном мозге три продольных магистрали:
1. Непарная вентральная спинномозговая артерия – лежит вместе с одноименной артерией в вентральной срединной щели, отдает ветви в серое мозговое вещество,
2. Парные спинномозговые дорсальные артерии – лежат вдоль дорсальных корешков нервов, а соответствующие вены – вдоль вентральных корешков. Все три артериальные магистрали анастомозируют межу собой в каждом сегменте, формируя сосудистый венец. От него отходят ветви в белое мозговое вещество, соединяясь внутри мозга с артериями серого вещества. Из вен кровь оттекает в венозные сплетения и парный позвоночный вентральный синус. Он лежит в эпидуральном пространстве и соединяется с сегментными венами туловища.
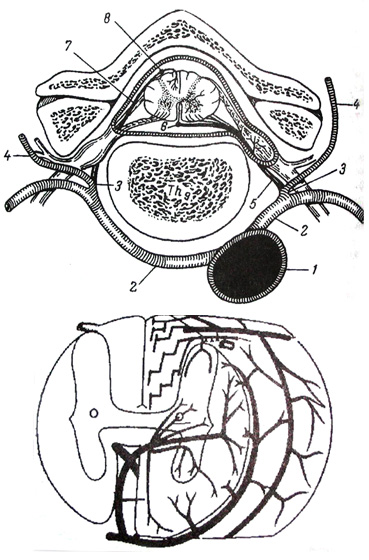 1. Аорта
1. Аорта
2. Межреберные артерии
3. Дорсальная ветвь ветвь
4. Мышечно - кожная ветвь
5. Спинальная ветвь
6. Вентральная радикуло-медулярная артерия(место перехода в вентральнуюспинальную артерию)
7. Дорсальная радикуло - медуллярная артерия 8 дорсальная спинальная артерия.
Сулько-комисуральная артерия
У собак отсутствует артерия Адамкевича данная артерия имеется только у людей и приматов это является существенным различием в кровоснабжении спинного мозга
- Периферические нервы
- Головной мозг
1. Продолговатый мозг
2. Мост мозга
3. Средний мозг
4. Мозжечок
5. Промежуточный мозг
6. Кора большого мозга
- Ликвороциркуляция
- Кровоснабжение центральной нервной системы.
Интерактивные функции спинного мозга (MEDULLA SPINALIS)
Спинной мозг– самый каудальный отдел центральной нервной системы
Особенностью спинного мозга является четко выраженное сегментарное строение
Общее количество сегментов соответствует числу метамеров тела (метамер - это сегмент, который получает чувствительные волокна от одной отдельной пары дорсальных корешков). Кожная область, которая иннервируется данными чувствительными нервами, называется дерматомом.
От каждого сегмента отходят одна пара передних, или вентральных, корешков, и одна пара задних, или дорсальных, корешков. Функциональная значимость данных корешков различна. Белл и Мажанди установили, что вентральные корешки состоят из эфферентных, “двигательных” волокон, дорсальные - из афферентных, “чувствительных” волокон. Установленная закономерность определяется как “закон Белла-Мажанди”. Передние и задние корешки кнаружи от спинномозговых узлов в межпозвоночном отверстии соединяются в смешанный спинномозговой нерв, который при выходе из позвоночника делится на дорсальную , вентральную ветви и ветвь, направляющуюся к симпатическому стволу (rammus communicans).
Спинной мозг делится на отделы:
- шейный,
- грудной,
- поясничный,
- крестцовый хвостовой.
На протяжении спинного мозга имеются два утолщения веретенообразной формы. Шейное утолщение образуется четырьмя шейными сегментами и первыми грудными, поясничное утолщение образуется четырьмя поясничными сегментами и тремя крестцовыми сегментами. Данные утолщения соответствуют местам выхода из спинного мозга корешков нервов для передних и задних конечностей. Спинной мозг состоит из белого вещества, образованного из миелиновых нервных волокон, и серого вещества, содержащего нервные клетки. Серое вещество спинного мозга заложено внутри и со всех сторон окружено белым веществом. Столб серого вещества образует три выступа: вентральный, дорсальный и боковой, которые на поперечных срезах мозга имеют форму рога. Соответственно различают вентральный, дорсальный и боковой. Вид серого вещества на поперечном срезе, как считают многие исследователи, напоминает букву “Н” или бабочку с раскрытыми крыльями. вентральный рог имеет округлую форму и содержит клетки, дающие начало передним двигательным корешкам. Дорсальный рог уже и длиннее переднего, содержит клетки, дающие начало задним чувствительным корешкам. Боковой рог определяется на протяжении последнего шейного, всех грудных и I-II поясничных сегментов спинного мозга. Боковой рог образует небольшой треугольный выступ латерального края серого вещества. В нем находятся мелкие по величине нейроны, аксоны которых выходят из спинного мозга вместе с вентральными и отчастидорсальными нервными корешками. Нейронный состав серого вещества спинного мозга сложен.
Различают следующие виды нейронов:
- Эфферентные нейроны, которые подразделяются на альфа-мотонейроны и гамма-мотонейроны.
- Преганглионарные нейроны. Их аксоны образуют преганглионарные нервные волокна, направляющиеся к ганглиям пограничного нервного столба.
- Интернейроны – это самая большая группа нейронов, которая участвует в интеграции процессов возбуждения и торможения. Отростки данных нейронов в основном обеспечивают внутрисегментарные и межсегментарные связи.
- Афферентные нейроны. Нейроны данного типа имеют один аксон, который Т-образно разделяется. Одна ветвь такого нейрона передает возбуждение от рецептора к телу нервной клетки, другая ветвь обеспечивает проведение возбуждения от тела спинального нейрона к другим спинальным нейронам. Эфферентные нейроны расположены в переднем роге и являются моторными центрами спинного мозга. Афферентные нейроны расположены в заднем роге и являются центрами, воспринимающими афферентацию от рецепторов. Нервные клетки бокового рога являются вегетативными центрами спинного мозга.
В 1925 году американский анатом Б. Рексед предложил серое вещество спинного мозга разделить на десять пластин или слоев, поверхности которых располагаются параллельно дорсальной или вентральной поверхности спинного мозга. Пластины обозначаются римскими цифрами. По существу, Рексед предложил функциональную топографию нейронов спинного мозга, представленную десятью пластинами. Они следующие: I-IV пластины образуют головку дорсального рога серого вещества – это первичная сенсорная область. В эту область проецируется большая часть афферентных волокон от туловища и конечностей. Отсюда берут начало несколько трактов спинного мозга, идущих в головной мозг. V-VI пластины образуют шейку дорсального рога. Здесь заканчиваются волокна от сенсомоторной области коры мозга и волокна, несущие проприоцептивную чувствительность от туловища и конечностей. VII пластина представляет область окончания проприоспинальных и висцеральных связей, а также афферентных и эфферентных связей спинного мозга с мозжечком и средним мозгом. Та часть VII пластины, которая находится в области вентрального рога, содержит клетки Реншоу. VIII пластина характеризуется бульбоспинальными и проприоспинальными связями. X пластина является первичной моторной областью и состоит из мотонейронов. Мотонейроны этой области объединены в функциональные группы, пулы (англ. – совокупность). X пластина занимает пространство вокруг спинномозгового канала и состоит из нейронов, клеток глии и комиссуральных волокон.
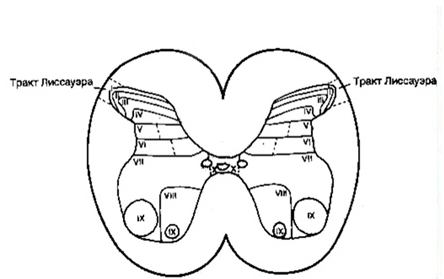
Пластины Рекседа
- Пластина I представляет собой самый поверхностный слой дорсального рога, ее еще называют краевым слоем. Он содержит большие плоские "маргиальные клетки" и нейроны промежуточного размера.
- Пластину II называют "желатинозной" из-за ее желатиноподобного вида на свежем срезе спинного мозга. В ее состав входят мелкие плотно расположенные клетки.
- Пластина III содержит крупные рыхло располагающиеся клетки.
- Пластина IV, самая толстая из расположенных в заднем роге, скомпанована из больших нейронов с дендритами, распространяющимися в другие пластины. Вместе пластины III и IV образуют собственное ядро (nucleus proprius).
- Пластина V состоит из мелких нейронов.
- Пластина VI локализуется в самом основании дорсального рога и прослеживается только в зонах утолщения спинного мозга (шейный и поясничный отделы). Весь дорсальныйрог сформирован пластинами I-VI.
- Пластина VII занимает неправильной формы область в центре серого вещества спинного мозга.
- Пластина VIII охватывает внутреннюю половину переднего рога в области шейного и поясничного его утолщений.
- Пластина IX соответствует расположению группы двигательных нейронов в вентральном роге, а
- Пластина Х окружает центральный канал. Таким образом, вентральный рог сформирован пластинами VII-X.
Белое вещество спинного мозга состоит из нервных волокон, которые делятся на эндогенные, или собственные, волокна, и экзогенные, или инородные. К эндогенным относятся волокна, берущие начало в спинном мозге; они могут быть длинными и короткими. Длинные направляются в головной мозг, короткие образуют межсегментарные связи.
Основными длинными эндогенными волокнами, или пучками, которые идут в восходящем направлении, являются следующие:
- Пучок Голля. Данный путь несет волокна от нижних конечностей и нижних отделов туловища.
- Пучок Бурдаха несет волокна от передних конечностей и передней половины туловища. Данные пучки занимают дорсальные канатики спинного мозга и заканчиваются в области продолговатого мозга.
- В боковых столбах спинного мозга проходит дорсолатеральный путь, проводящий болевую и температурную афферентацию.
- Прямой мозжечковый пучок, или пучок Флексига. Данный путь берет начало в клетках заднего рога и заканчивается на структурах мозжечка.
- Перекрещенный мозжечковый пучок Говерса. Берет начало из клеток заднего рога противоположной стороны, часть волокон пучка Говерса оканчивается в мозжечке (tr. spino-cerebellaris), в ядрах продолговатого мозга (tr. spino-bulbaris), в буграх четверохолмия (tr. spino-tectalis), зрительном бугре (tr. spino-talamicus lаt.).
- Спинно-оливарный пучок проходит на границе вентрального и бокового столбов. Данный пучок берет начало из клеток дорсального рога и оканчивается в районе олив продолговатого мозга.
Из пучков, идущих в нисходящем направлении, следует отметить:
- Пирамидный путь (tr. cortico-spinalis), который после перекреста волокон в продолговатом мозге делится на два пучка. Один из них идет в боковом столбе противоположной стороны спинного мозга (перекрещивающийся пирамидный путь) и заканчивается в клетках переднего рога своей стороны. Другой пирамидный пучок идет в переднем столбе той же стороны спинного мозга и заканчивается в клетках переднего рога противоположной стороны (прямой пирамидный путь).
- Пучок Монакова (tr. rubro-spinalis) берет начало в красных ядрах среднего мозга, по выходе из которых перекрещивается (перекрест Фореля) и заканчивается в клетках вентрального рога.
- Ретикуло-спинальный путь (tr. reticulo-spinalis) происходит от ретикулярной формации противоположной или своей стороны и заканчивается в клетках вентрального рога.
- Вестибуло-спинальный пучок (преддверно-спинномозговой путь, tr. vestibulo-spinalis) берет начало от клеток ядра Дейтерса и заканчивается в клетках вентрального рога.
- Пучок Гельвега (tr. praeolivaris) берет начало из области покрышки и заканчивается в клетках вентрального рога шейного отдела спинного мозга.
- Задний продольный пучок (fasc. longitudinalis dorsalis) начинается от различных клеток мозгового ствола и заканчивается в клетках вентрального рога
- Предтыльный пучок (tr. tecto-spinalis) берет начало в буграх четверохолмия, образует перекрест и оканчивается в клетках вентрального рога.
- Fasc. praepyramidalis Thomas начинается в ретикулярной формации ствола и оканчивается в клетках вентрального рога шейного отдела спинного мозга.
Система восходящих проводящих путей осуществляет функцию проведения импульсов от рецепторов, которые воспринимают информацию из внешнего мира и внутренней среды организма. В зависимости от вида чувствительности, которую они проводят, восходящие проводники делятся на пути экстеро-, проприо- и интероцептивной чувствительности. Система нисходящих проводящих путей осуществляет функцию проведения импульсов от различных отделов головного мозга к двигательным ядрам (клеткам) спинного мозга. В функциональном отношении нисходящие проводники могут быть охарактеризованы, в основном, как система волокон, осуществляющих двигательную функцию. Следует отметить, что в последние годы выявлена возможность проведения по данной системе афферентации к таким центрам продолговатого мозга, как дыхательный, вазомоторный и пищеварительные [Меркулова Н.А., Инюшкин А.Н., Беляков В.И., Зайнулин Р.А. и др.
Рефлекторные функции спинного мозга
Изучение и анализ рефлекторных функций спинного мозга следует проводить на “спинальном животном”. “Спинальное животное” - это животное, у которого среди всех отделов центральной нервной системы сохранен только спинной мозг. Для “приготовления” “спинального животного” необходимо произвести перерезку мозга каудальнее продолговатого мозга. У всех позвоночных животных перерезка мозга под продолговатым полностью или в значительной степени подавляет рефлекторную деятельность спинного мозга. Состояние подавления рефлекторной деятельности мозга известно под названием шока (означает удар, сотрясение).
Это название было дано английским ученым Маршал Холлом (1835 г.). Явление шока обнаруживается у различных позвоночных животных в различной степени. Чем выше эволюционная ступень, которую животное занимает, тем продолжительнее состояние шока. У человека и высших обезьян после перерезки спинного мозга состояние шока продолжается в течение нескольких лет, а иногда навсегда утрачивается способность к рефлекторной деятельности [Беритов, 1948]. У кошек и собак рефлекторная деятельность восстанавливается через несколько дней или недель; у кроликов - через несколько часов; у амфибий - через одну-десять минут. У низших млекопитающих животных, у всех низших позвоночных состояние шока наблюдается, главным образом, в отношении скелетной мускулатуры. Из вегетативных органов шоку подвергаются только органы сосудистой системы.
Но у высших позвоночных животных, шок в одинаковой степени захватывает как соматическую, так и вегетативную системы: наблюдается паралич двигательных рефлекторных реакций, остановка дыхания, резкое понижение артериального давления, “паралич” кишечника, мочевого пузыря, понижение температуры тела. Состояние шока при перерезке мозга проявляется не в одинаковой степени во всех элементах спинного мозга. Анализ биоэлектрической активности нейронов спинного мозга выявил, что состоянию шока после перерезки спинного мозга подвергаются, главным образом, моторные нейроны. Следует отметить, что угнетенное состояние нервных элементов более выражено в каудальном направлении, чем в краниальном. Угнетенное состояние рефлекторной деятельности каждого отдела спинного мозга зависит от его близости к разрезу мозга. Например, если спинной мозг перерезан в шейном отделе, то состояние шока проявляется на передних (верхних) конечностях сильнее, чем на задних (нижних). Относительно природы шока имеется ряд мнений. Впервые немецкий физиолог Гольц (1896 г.) высказал мнение, что причиной шока является торможение нервных элементов спинного мозга, вызванное травмой. Однако английский физиолог Шеррингтон (1906 г.), тщательно изучивший явление спинального шока, показал, что шок нельзя объяснить торможением структур спинного мозга.
В пользу мнения Шеррингтона можно привести следующие факты:
- Если бы шок был торможением структур спинного мозга, то он обнаруживался бы в краниальном отделе с такой же силой, что и в каудальном.
- После перерезки спинного мозга под продолговатым мозгом развивается яркая картина спинального шока.
Если после того, как восстановится рефлекторная деятельность спинного мозга, вновь перерезать спинной мозг ниже прежнего уровня перерезки, то явления спинального шока не проявляются. Учитывая два приведенных факта, неодинаковую продолжительность шока у различных представителей животного мира, а также электрофизиологические исследования спинального шока, в последние годы сформулирован современный взгляд на природу спинального шока. Сущность его заключается в следующем: одним из главных факторов, вызывающих явление шока при перерезке спинного мозга, является разрыв длинных путей, нисходящих из головного мозга, что приводит к внезапному прекращению многообразной афферентации из структур головного мозга на центры спинного мозга. Прекращение потока афферентации из головного мозга расстраивает (угнетает) рефлекторную деятельность спинного мозга. В происхождении шока некоторую роль играет и другой фактор. Перерезка мозга вызывает достаточно длительное механическое раздражение спинного мозга. Подвергаются раздражению не только нервные клетки, но и восходящие и нисходящие пути, что, в конечном итоге, приводит к угнетению рефлекторной деятельности. После того как исчезнут явления спинального шока, можно наблюдать следующие рефлексы спинного мозга: защитные рефлексы, рефлексы на растяжение, рефлексы мышц-антогонистов, висцеромоторные и вегетативные рефлексы. Защитные рефлексы у лягушки обычно проявляются в отдергивании лапки при слабом раздражении кожных рецепторов, при более сильном болевом раздражении можно наблюдать “убегание” животного. Рефлексы растяжения проявляются в укорочении мышцы при ее растяжении. Рефлексы мышц-антагонистов лежат в основе локомоторных актов ходьбы и бега. Висцеральные рефлексы проявляются при раздражении афферентных волокон внутренних органов. Вегетативные рефлексы проявляются, в основном, при возбуждении преганглионарных симпатических волокон в ответ на возбуждение симпатических и соматических чувствительных клеток. При поражении верхнего шейного отдела спинного мозга возникает паралич шейных мышц, диафрагмы, анестезия в области шеи и затылка. При поражении шейного утолщения развивается паралич передних конечностей, их анестезия. При поражении грудного отдела наступает парез (частичный паралич) мышц спины, грудной или брюшной стенки с сегментарной анестезией. При поражении поясничного утолщения наблюдается паралич задних конечностей, анестезия в нижних конечностях, расстройство тазовых органов. Поражение конуса (конский хвост; сегменты СI-III) вызывает анестезию в области промежности и расстройство тазовых органов.
В спинном мозге расположены следующие важнейшие вегетативные центры:
- В боковых рогах грудного отдела спинного мозга находятся вазомоторные центры и центры потовых желез.
- На уровне краниальныхпоясничных сегментов и в третьем, четвертом и пятом крестцовых сегментах заложены центры мочеиспускания и дефекации.
- На уровне крестцовых сегментов находятся центры эрекции и эякуляции.
- На уровне VII шейного - IV поясничного сегментов расположены центры симпатической нервной системы.
- На уровне 1- III - сакральных сегментов спинного мозга находятся центры парасимпатической нервной системы. При поражении центров мочеиспускания наступает паралич сфинктера и детрузора, вследствие чего моча постоянно по каплям выделяется наружу. При поражении центров дефекации наступает недержание кала и газов, отсутствует анальный рефлекс. При поражении центров половых рефлексов нарушаются эрекция и эякуляция.
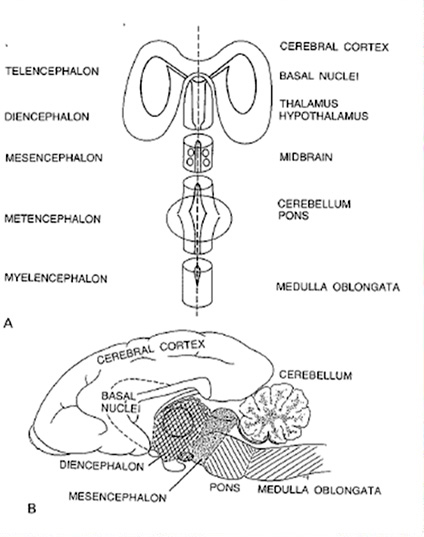
Сегментарное строение головного мозга
Интегративные функции продолговатого мозга
Продолговатый мозг - самая нижняя часть головного мозга, расположен между варолиевым мостом и спинным мозгом. Продолговатый мозг имеет важное функциональное значение.
Его основные функции следующие:
- Проводниковая функция.
- Продолговатый мозг включает ряд важных, жизненно необходимых рефлекторных центров.
- В продолговатом мозге расположены центры некоторых черепно-мозговых нервов.
- Нервные центры продолговатого мозга принимают участие в регуляции мышечного тонуса и некоторых установочных рефлексов.
- Продолговатый мозг содержит ретикулярную формацию.
Характеризуя проводниковую функцию продолговатого мозга, следует отметить, что в нем располагаются волокна, несущие импульсы от различных структур головного мозга к периферии и от периферии к структурам головного мозга. В продолговатом мозге имеются волокна ретикулярной формации. Пути восходящего и нисходящего направлений описаны выше Из жизненно необходимых рефлекторных центров прежде всего следует назвать дыхательный и сосудодвигательный (вазомоторный). Благодаря исследованиям нескольких поколений отечественных и зарубежных физиологов в настоящее время общепринятым стало положение о том, что ведущая роль в регуляции дыхания принадлежит структурам продолговатого мозга. Дыхательный центр рассматривается как совокупность дыхательных нейронов, активность которых синхронна с фазами дыхательного цикла. В соответствии с характером паттерна активности дыхательные нейроны подразделяются на шесть основных типов (Bianchi et al., 1995): ранние инспираторные, инспираторные с нарастающим паттерном активности, поздние инспираторные, постинспираторные, экспираторные с нарастающим паттерном активности,преинспираторные.
Дыхательные нейроны сосредоточены, главным образом, в пяти функционально различных областях дыхательного центра:
- Дорсальная дыхательная группа нейронов, расположенная в вентролатеральном отделе ядра солитарного тракта.
- Ростральная (инспираторная) часть вентральной дыхательной группы, находящаяся в области n. Ambiguus.
- Каудальная (экспираторная) часть вентральной дыхательной группы, находящаяся в области n. Retroambigualis.
- Комплекс пре-Бетцингера, расположенный в ростральной части n. ambiguus и вентролатеральной области ретикулярной формации каудальнее n. retrofacialis и ростральнее n. lateralis reticularis (3 мм ростральнее obex, 3,2-4 мм латеральнее средней линии). Данный комплекс содержит уникальное разнообразие типов дыхательных нейронов. Здесь присутствет большое количество проприобульбарных нейронов, имеются бульбоспинальные нейроны и краниальные мотонейроны, нейроны, участвующие генерации ритма дыхания (преинспираторные и нейроны с пейсмекерными свойствами), выявлены пре- и постинспираторные нейроны.
- Комплекс Бетцингера. Данный отдел дыхательного центра расположен в области n. retrofacialis.
Большинство клеток комплекса Бетцингера относится к экспираторным нейронам. Такие нейроны образуют моносинаптические ингибиторные проекции в направлении бульбоспинальных инспираторных нейронов дорсальной и вентральной дыхательной группы, каудальной группы ядер шва. В комплексе Бетцингера имеются также пейсмекерные нейроны. В начале XIX века Флуранс (Flourans, 1824) высказал мнение о том, что в регуляции кровообращения наиболее важную роль играет продолговатый мозг. В 1853 году Бюдж (Budg), а затем в 1855 году Шифф (Shiff) установили, что перерезка спинного мозга под продолговатым мозгом вызывает резкое падение артериального давления. В связи с этим они сделали вывод, что центр, регулирующий величину артериального давления, находится в продолговатом мозге. Наиболее фундаментальные исследования по анализу местоположения сердечно-сосудистого центра были проведены Я.А. Дедюлиным (1868) на холоднокровных животных и Диттмаром (Dittmar, 1873) и Ф.В. Овсянниковым (1871) на теплокровных животных.
Особого внимания заслуживают работы Ф.В. Овсянникова. Он установил, что в области, верхняя граница которой расположена на 1-2 мм каудальнее четверохолмия, а нижняя - на 4-5 мм ростральнее писчего пера, расположен центр, регулирующий деятельность сердечно-сосудистой системы. При разрушении этой области происходит необратимое выраженное понижение артериального давления. Дальнейшие исследования локализации вазомоторного центра показали следующее. Ляфон (Laffont, 1880) установил, что при локальном механическом раздражении различных участков дна четвертого желудочка продолговатого мозга могут возникать прессорные и депрессорные реакции. С.А. Бруштейн (1901) показал, что вазомоторный центр, вызывающий прессорные и депрессорные реакции, расположен под дном ромбовидной ямки, в ее средней и нижней трети, в ретикулярной формации продолговатого мозга. Представление о локализации в продолговатом мозге двух вазомоторных центров (прессорного центра, повышающего давление, и депрессорного центра, понижающего давление) получило развитие в работах Бейлиса (1893-1923). Портер (Porter, 1915) считал, что в продолговатом мозге имеются два центра: вазотонический, осуществляющий контроль сосудистого тонуса, и вазорефлекторный, интегрирующий сердечно-сосудистые рефлекторные реакции. Рэнсон и Биллингслей (Ranson, Billingsley, 1916) высказали мнение, что прессорный центр расположен в области forea inferior, у вершины ala cineria, а депрессорный центр находится в области area postrema, несколько латеральнее obex. Различная локализация прессорного и депрессорного центров показана в работах многих исследователей (Scott, Roberts, 1923; Wang, Ranson, 1939; Склярский, 1941 и др.). В 1946 году Александер (Alexander) высказал мнение о том, что в продолговатом мозге имеется только один центр - вазомоторный. Он представляет собой единое функциональное образование с эфферентными путями, которые идут в составе дорсолатеральных канатиков спинного мозга.
Изложенные выше представления о четкой дифференциации прессорных и депрессорных структур продолговатого мозга не получили подтверждения в работах многих исследователей. В настоящее время сложилось мнение, что в продолговатом мозге расположен основной сосудодвигательный центр, который поддерживает тонус сосудов и обеспечивает рефлекторную регуляцию артериального давления. Это мнение впервые было обосновано Ф.В. Овсянниковым в 1871 году. Он показал, что разрушение только продолговатого мозга вызывает необратимое “катастрофическое” падение артериального давления. Но четкая анатомическая локализация сосудодвигательного центра в продолговатом мозгу к настоящему моменту так и не установлена. Возможно, нейроны, регулирующие уровень артериального давления, диффузно расположены в продолговатом мозгу.
Также существует мнение, что сосудодвигательный центр состоит из трех основных типов нейронов: прессорных, депрессорных и кардиоингибирующих. Прессорные нейроны (группа, зона) повышают артериальное давление в результате увеличения периферического сопротивления сосудов и увеличения сердечного выброса; депрессорные нейроны (группа, зона) понижают артериальное давление, вызывая торможение тонических разрядов вазоконстрикторов; кардиоингибирующие нейроны (группа, зона) уменьшают величину сердечного выброса, возбуждая сердечный центр блуждающего нерва. В структурах продолговатого мозга расположены: пищеварительный центр, состоящий из нескольких компонентов, центры слюноотделения, потоотделения, центры защитных дыхательных рефлексов, рвоты, регуляции углеводного обмена.
Относительно центра углеводного обмена следует отметить, что впервые в 1849 году французский физиолог Клод Бернар произвел опыт, получивший название “сахарного укола”, и этим положил начало исследованиям бульбарной регуляции углеводного обмена. Было обнаружено, что раздражение заднего отдела дорсального ядра блуждающего нерва вызывает гипергликемию и гликозурию, а раздражение переднего отдела ядра приводит к снижению сахара в крови и моче. На этом основании было высказано мнение, что в продолговатом мозге имеются два центра, один из которых повышает содержание сахара в крови и моче, другой - понижает. В продолговатом мозге находится бульбарный отдел парасимпатической нервной системы. Он представлен клеточными группами ядер лицевого, подъязычного, языкоглоточного и блуждающего нервов. Парасимпатические волокна лицевого нерва иннервируют слезную железу, подчелюстную и подъязычную слюнные железы. Парасимпатические волокна блуждающего нерва иннервируют щитовидную и вилочковую железы, бронхи, легкие, сердце, пищевод, желудок, тонкие и толстые кишки до поперечной ободочной кишки включительно, печень и почки. Парасимпатические волокна языкоглоточного нерва иннервируют подчелюстную и околоушную слюнные железы.
В продолговатом мозге заложены ядра многих черепно-мозговых нервов. VIII пара - слуховой нерв (n. acusticus). Ядра этого нерва лежат на дне ромбовидной ямки. Они состоят из двух различных по функции корешков: n. cochlearis, нерв улитки, является слуховым нервом; n. vestibularis, вестибулярный нерв, является центром проприоцептивной чувствительности, регулирующим равновесие тела и координацию движений. IX пара - языкоглоточный нерв (n. glossopharyngeus) - смешанный нерв, состоящий из двигательных и чувствительных (главным образом, вкусовых) волокон.
Двигательное ядро данного нерва находится в продолговатом мозге. Вкусовые волокна берут начало из ganglion jugulare et ganglion petrosum. Языкоглоточный нерв проводит вкусовую афферентацию с рецепторов слизистой оболочки задней трети языка и мягкого неба с его передними дужками. Х пара - блуждающий нерв, n. vagus, является смешанным нервом. Он содержит чувствительные и двигательные волокна. Двигательные волокна берут начало в дорсальном ядре (n. dorsalis) и вентральном ядре (n. ambiguus). Они иннервируют небные мышцы, шилоглоточную, мышцы гортани, а также все органы грудной и брюшной полостей. Чувствительные волокна блуждающего нерва берут начало от клеток ganglion jugulare et ganglion nodosum. Чувствительные волокна блуждающего нерва проводят афферентацию от рецепторов всех внутренних органов, а также от рецепторов кожи наружного слухового прохода и ушной раковины. XI пара - добавочный нерв, n. accessorius Willissii. Часть волокон этого нерва выходит из каудальной части продолговатого мозга. Большая часть клеток, дающая начало добавочному нерву, находится в шейном отделе спинного мозга. Другая часть клеток примыкает к двигательному ядру блуждающего нерва. Добавочный нерв состоит только из двигательных волокон. Он иннервирует две мышцы: m. sterno-cleido-mastoideus et m. trapezius. XII пара - подъязычный нерв, n. hypoglossus. Нерв содержит только двигательные волокна. Он иннервирует мускулатуру языка.
Одна из структур продолговатого мозга – парное ядро Дейтерса, которое наряду с красными ядрами и буграми четверохолмия принимает участие в регуляции тонуса мышц. Наиболее наглядно это участие проявляется в “децеребрационной ригидности”. Децеребрация - это отделение части головного мозга от другой. Децеребрация впервые была произведена в 1896 году английским физиологом Чарльзом Шеррингтоном. В случае “децеребрационной ригидности” перерезку головного мозга обычно производят между передними и задними буграми четверохолмия, реже между задними буграми четверохолмия и продолговатым мозгом. После операции по мере ослабления наркоза развивается децеребрационная ригидность. Она проявляется в том, что все конечности разгибаются и судорожно вытягиваются, голова и шея поднимаются вверх, вверх поднимается хвост и “выгибается” спина. Во время ригидного состояния сокращаются и мышцы-сгибатели, однако механическое действие разгибателей на суставы сильнее, чем сгибателей, поэтому сохраняется разгибательное положение. При этом следует отметить, что во время сильного ригидного состояния мышцы-сгибатели конечностей испытывают сильное тоническое торможение. С течением времени децеребрационный разгибательный тонус ослабевает и может смениться на общий сгибательный тонус. Механизм децеребрационной ригидности следующий: ядра Дейтерса продолговатого мозга находятся под постоянным тормозным влиянием красных ядер среднего мозга. Красные ядра не только оказывают постоянное тоническое, тормозное влияние на ядра Дейтерса, но и обеспечивают равномерность распределения афферентации между мышцами-сгибателями и мышцами-разгибателями. После отделения красных ядер от ядер Дейтерса прекращается тормозное и другие влияния красных ядер на ядра Дейтерса, что и приводит к развитию разгибательного гипертонуса. Тормозное влияние на ядра Дейтерса оказывает и мозжечок (через фастигиальное ядро), поэтому удаление мозжечка ведет к усилению децеребрационной ригидности. На децеребрационных животных можно наблюдать позные установочные рефлексы, фазные рефлексы чихания, “ходьбы”. В ретикулярной формации продолговатого мозга располагаются многие сложные центры.
Исследования показали, что определенные области продолговатого мозга влияют на мотонейроны спинного мозга. Эти бульбарные нейроны, в свою очередь, находятся под воздействием вышележащих областей мозга. В вентролатеральной части ретикулярной формации продолговатого мозга выявлена группа клеток, которая оказывает тормозящее влияние на спинальные рефлексы. В дорсальной части ретикулярной формации продолговатого мозга расположена группа клеток, которая обеспечивает осуществление спинальных рефлексов. Особого внимания заслуживает одно из ядер ретикулярной формации - гигантоклеточное ядро. Работы сотрудников кафедры физиологии человека и животных Самарского госуниверситета (Н.А. Меркуловой, А.Н. Инюшкина, В.И. Белякова, Р.А.Зайнулина) позволили сделать следующий вывод: респираторные влияния сенсомоторной коры мозга, мозжечка, а также структур экстрапирамидной системы реализуются через ретикулярное гигантоклеточное ядро. Данное ядро, с известной долей вероятности, можно рассматривать как коллектор многообразной афферентации, которая поступает к дыхательному центру от различных супрабульбарных отделов головного мозга.
Интегративные функции заднего мозга
Задний мозг состоит из двух отделов: варолиева моста и мозжечка.
Варолиев мост, или просто мост (pons), представляет собой со стороны основания мозга толстый белый вал, граничащий каудально с ростральным концом продолговатого мозга, а краниально – с ножками мозга.
В варолиевом мосту расположены ядра V-VIII пары черепно-мозговых нервов.
V пара - отводящий нерв (n. abducens), ядро этого нерва расположено в краниальном отделе варолиева моста. Этот нерв иннервирует только одну мышцу - мышцу, отводящую глаз кнаружи.
VI пара - тройничный нерв (n. trigeminus), который состоит из двигательных и чувствительных волокон. Чувствительные волокна иннервируют краниальный отдел головы, кожи лба и верхнего века, конъюнктиву глазного яблока, роговую оболочку и радужку, а также слизистую оболочку лобной пазухи и верхней части носа. Тройничный нерв иннервирует кожу, нижних частей носа, верхней челюсти и неба, а также верхние и нижние зубы, слизистую оболочку щек, нижней челюсти, дна полости рта, языка; снабжает вкусовыми волокнами передние две трети языка.
VII пара - лицевой нерв (n. facialis). Данный нерв иннервирует передней части морды мускулатуру. Раздражение вестибулярных ядер варолиева моста вызывает повышение артериального давления, увеличение периферического сопротивления сосудов и уменьшение сердечного выброса. Наряду с гемодинамическими изменениями при электростимуляции различных участков вестибулярных ядер моста отмечаются многообразные изменения дыхания: уменьшение или увеличение глубины дыхания, учащение или урежение дыхания. На сновании этих данных можно считать, что варолиев мост принимает участие в регуляции дыхания, сосудистого тонуса и деятельности сердца.
Мозжечок представляет собой вырост моста. Он появляется на ранних этапах филогенеза позвоночных. Мозжечок может быть различным по своей величине - от небольшого “комочка” до крупного образования. У некоторых рыб он достигает значительных размеров, но у амфибий и рептилий он мал. Значительное развитие мозжечок получает у млекопитающих животных и человека. О. Ларсел всю поверхность мозжечка разделяет на несколько отделов, главным образом, в зависимости от филогенетического возраста.
Эти отделы следующие:
- Архицеребеллум (древний мозжечок) представлен небольшой по величине клочково-узелковой долькой.
- Палеоцеребеллум (старый мозжечок) включает переднюю долю, участок червя, соответствующий передней доли, пирамиды, язычок, парафлокулярную долю.
- Неоцеребеллум (новый мозжечок) включает полушария и часть червя, которая расположена каудальнее участка червя, соответствующего передней доле.
Обращает на себя внимание строение коры полушарий мозжечка. Она имеет четко выраженное трехслойное строение.
Первый поверхностный слой - молекулярный. Состоит из клеток корзинчатой и звездчатой форм.
Второй слой - гранулярный - представлен клетками Пуркинье, которые встречаются только в мозжечке.
Третий слой - зернистый - состоит из зернистых клеток и клеток Гольджи.
По данным Фанарджяна, в коре мозжечка имеется пять типов клеток:
- клетки Пуркинье,
- корзинчатые клетки,
- звездчатые клетки,
- клетки Гольджи,
- зернистые клетки.
По данным Шмида, в коре полушарий мозжечка наряду с вышеназванными типами клеток имеется шестой тип клеток - клетки Лугаро.
Мозжечок имеет широко развитые связи, по существу, со всеми структурами головного мозга, а также со спинным мозгом.
Основные афферентные пути мозжечка следующие:
- Дорсальный спинно-мозжечковый тракт.
- Вентральный спинно-мозжечковый тракт, проводящий проприоцептивную афферентацию от задней части тела.
- Ростральный спинно-мозжечковый тракт, проводящий проприоцептивную афферентацию от передней части тела.
- Спинно-оливо-мозжечковый тракт.
- Церебро-мозжечковые связи. По данным связям афферентация поступает в мозжечок из “моторной” области коры больших полушарий головного мозга.
- Кортико-ретикуло-мозжечковый путь.
- Оливо-мозжечковый тракт. Данный путь проводит афферентацию из области олив в мозжечок.
- Вестибуло-мозжечковый путь передает афферентацию от вестибулярных ядер в мозжечок.
- Рубро-мозжечковые связи, передающие афферентацию из красных ядер в мозжечок.
- Ретикуло-мозжечковые связи проводят афферентацию к коре полушарий мозжечка от латерального, парамедиального ядер продолговатого мозга, от ядра покрышки варолиева моста, от ретикулярного гигантоклеточного ядра.
- Выявлены проводящие пути от структур базальных ганглиев к мозжечку.
Все афферентные пути оканчиваются в виде трех видов волокон. Мшистые волокна идут от ядер моста и оканчиваются в зернистом слое коры мозжечка. Лиановидные, или лазающие, волокна идут от нижних олив. Данные волокна представляют уникальный компонент организации коры мозжечка. Одно лиановидное волокно устанавливает синаптический контакт только с одной клеткой Пуркинье.
На уровне слоя клеток Пуркинье данные волокна теряют миелин и проходят параллельно телу и дендритам клеток Пуркинье. Лиановидные волокна, проходя через зернистый слой, отдают коллатерали на синапсы дендритов зернистых клеток, соме клеток Гольджи, клеток Лугаро. Третья афферентная система - моноаминоэргические связи. Эта система включает норадренэргические, серотонинэргические и дофаминэргические волокна. Источником норадренэргических волокон является голубое пятно. Волокна от голубого пятна идут ко всем ядрам мозжечка, проходят через зернистый слой, а затем оплетают клетки Пуркинье и вступают в молекулярный слой. Дофаминэргические волокна поступают в мозжечок из области покрышки среднего мозга. Эти волокна образуют синаптические контакты с клетками Пуркинье и зернистыми клетками. Источником серотонинэргических волокон являются ядра продолговатого, среднего мозга и моста.
Основные эфферентные пути мозжечка следующие. Установлено, что аксоны клеток Пуркинье, являющиеся тормозными нейронами, составляют единственный эфферентный путь. Но волокна, составляющие этот эфферентный путь, осуществляют проведение преимущественно, если не ислючительно, тормозящих влияний к многочисленным структурам центральной нервной системы: спинному мозгу, к ядрам продолговатого, среднего и промежуточного мозга, центрам экстрапирамидной системы, “моторной” области коры головного мозга. Следует отметить, что моховидные волокна проводят афферентацию возбуждающего характера. Лиановидные волокна, опосредованные через нейроны Пуркинье, отчасти через корзинчатые и звездчатые нейроны, проводят афферентацию тормозящего характера. Таким образом, мозжечок может оказывать разнообразные влияния - возбуждающие и тормозящие на различные отделы центральной нервной системы.
Важную функциональную роль играют ядра мозжечка.
В белом веществе мозжечка расположены следующие парные ядра:
ядра шатра, пробковидные, шаровидные и зубчатые ядра.
Отмеченные ядра имеют связи с многочисленными структурами центральной нервной системы (спинным мозгом, продолговатым мозгом, мостом, средним и промежуточным мозгом, моторной зоной коры больших полушарий). ля изучения функций мозжечка используются различные методы.
Основными являются: метод клинических наблюдений, метод экстирпации (удаления), раздражения, электрофизиологические методы. Удаление мозжечка позволило прежде всего выявить его особую роль в интеграции информации, необходимой для регуляции двигательных реакций (Лучиани, 1893; Левандовский, 1907; Орбели, 1935; Алексанян, 1948; Карамян, 1956, 1970; Моруцци, 1958; Аршавский, 1976; Григорян, 1976 и др.).
Установлены основные функции мозжечка в регуляции двигательной активности:
- регуляция позы и мышечного тонуса;
- коррекция медленных целенаправленных движений;
- обеспечение выполнения быстрых целенаправленных движений.
После удаления мозжечка выявляются следующие нарушения (симптомы нарушения функции мозжечка):
- Асинергия - отсутствие посылки должного количества импульсов к различным мышцам, выполняющим движения. Это приводит к тому, что движения выполняются или в избыточном, или недостаточном объеме. Наблюдается неправильная походка с широко расставленными ногами и избыточным объемом двигательных реакций. Данный симптом впервые описан Бабинским в 1899 году.
- Астазия - колебательные движения головы и туловища. Тремор усиливается во время двигательной активности, в состоянии покоя тремор исчезает.
- Атаксия - нарушение величины, силы, скорости, направления двигательных реакций. Движения утрачивают плавность и стабильность, развивается дисметрия (неправильная оценка расстояния).
- Гипотония - понижение мышечного тонуса. Чаще развиваются волнообразные изменения тонуса: гипотония сменяется повышением тонуса мышц, в дальнейшем снова происходит понижение тонуса мышц и так далее.
- Нистагм - непроизвольные движения глазных яблок.
- Головокружение.
- Астения - быстрая утомляемость.
Многообразие симптомов, которые развиваются после удаления мозжечка, по-видимому, объясняется обилием эфферентных связей данной структуры с различными отделами центральной нервной системы. Возможно, мозжечок согласует работу различных структур в единую систему, которая определяет адекватность и совершенство двигательных реакций. Имеются и другие мнения о значении мозжечка в регуляции двигательных реакций. Так, Виннер (1961) считает, что мозжечок играет роль системы, которая предупреждает возникновение колебательных режимов при выполнении движений. Рух (1951) рассматривает мозжечок как своеобразный блок, который обеспечивает сравнение команд, посылаемых корковыми центрами регуляции движений с реальным ходом их выполнения. На основании такого сравнения мозжечок коррегирует работу исполнительных двигательных центров. Брайтенберг (1967) считает, что мозжечок осуществляет точное измерение временных интервалов между афферентными сигналами.
С 30-х годов ХХ века были предприняты систематические исследования Л.А. Орбели, посвященные значению мозжечка в регуляции вегетативных функций. Установлена роль мозжечка в регуляции многих вегетативных функций: пищеварения, дыхания, сосудистого тонуса, деятельности сердца, терморегуляции, обмене веществ и других.
На кафедре физиологии человека и животных Самарского госуниверситета были проведены исследования по анализу значимости мозжечка в регуляции дыхания (Н.А. Меркулова, А.Н. Инюшкин, В.И. Беляков). Сравнительный анализ респираторных реакций, вызванных электростимуляцией различных участков структур мозжечка, позволил выявить угнетение ритмогенерирующей функции дыхательного центра. Установлено, что наиболее активные в отношении регуляции дыхания участки мозжечка у крысы топически перекрываются с областями моторного представительства вибриссного аппарата и передних конечностей. В механизме реализации респираторных влияний мозжечка участвует ГАМК-ергическая нейромедиаторная система. “Мишенями” реализации дыхательных реакций мозжечка являются амбигуальное и ретикулярное гигантоклеточное ядра продолговатого мозга.
Интегративные функции среднего мозга
В состав среднего мозга входят ножки мозга и четверохолмия. Ножка мозга представляет собой массивный тяж продольных нервных волокон, идущий от переднего края варолиева моста в массу полушария головного мозга. Вследствие расхождения ножек между ними образуется ямка, дно которой усеяно многочисленными отверстиями, служащими для прохождения сосудов с основания мозга вглубь полушарий головного мозга.
Дорсальная часть среднего мозга образована пластинкой четверохолмия, лежащей над сильвиевым водопроводом. Пластинка имеет четыре возвышения: два передних образуют переднее двухолмие (передние бугры четверохолмия), два задних возвышения - заднее двухолмие (задние бугры четверохолмия).
На уровне передних бугров четверохолмия, на дне сильвиева водопровода лежит ядро III пары черепно-мозговых нервов глазодвигательных нервов (n. oculomotorius).
На уровне задних бугров четверохолмия, также на дне сильвиева водопровода лежит ядро IV пары черепномозговых нервов блоковых нервов (n. trochlearis). В ножке мозга различают основание и покрышку. Границу между основанием и покрышкой образует черное вещество Земмеринга (substantia nigra Soemmeringi). В покрышке мозговой ножки лежит красное ядро (n. ruber).
Анализ морфологических особенностей среднего мозга позволяет выделить следующие основные структуры, обеспечивающие многие важные функции: ядро глазодвигательного нерва, ядро блокового нерва, красное ядро, черная субстанция.
Давая общую характеристику функциям среднего мозга, следует отметить:
- проводниковую функцию;
- наличие в среднем мозге центров многих рефлекторных реакций,особенно локомоторных.
 Рис. 1. Спинной мозг таксы в позвоночном канале.
Рис. 1. Спинной мозг таксы в позвоночном канале.



